1.まえがき
昨年8月2-8日に北海道大学で第3回バイオメカニックス世界会議が開催されたが、その席上、ケンブリッジ大学のPedley教授が、Lighthill教授の追悼のセミナーをされた。Lighthill教授は7月17日に亡くなられたとの事であった。Nature(1)や日本流体力学会誌ながれ(2)に、追悼文が相次いでいる。Lighthill教授は、流体力学における今世紀最大の学者の一人であり、数多くの先駆的な研究によって流体力学の分野を創始された。その一つに生物流体力学(3)があり、鞭毛による微生物の運動、魚の推進、鳥の飛行、昆虫の飛翔の研究を基礎づけられた。
ここでは、水中で運動する微生物や魚の運動に関する基礎的研究について述べ、現在、国内で行われている水棲生物の研究について言及する。最近、水棲生物や昆虫に関する流体力学的研究が、活発になっていると感じられる。
2.鞭毛による推進機構(抵抗力推進)
微生物には、ミリサイズの線虫から、ミクロンサイズの細菌などがあるが、推進速度のレイノルズ数が1以下と非常に小さいことから、推進機構には、抵抗力が利用されている。
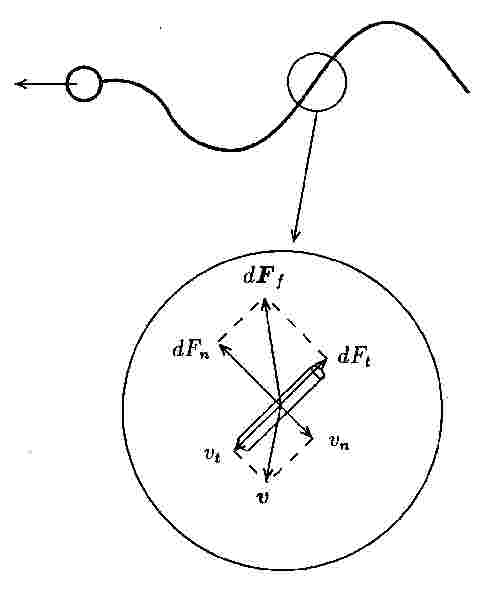
Grayら(4)は、鞭毛を屈曲して水中を移動するウニの精子の推進機構を解析したが、この手法は抵抗力理論に分類され、簡単に意味のある結果を得ることが出来る。この理論は、静止流体中を運動する鞭毛の円柱状の要素が受ける抵抗力を、図1に示すように、速度の接線成分および法線成分に対する抵抗係数を用いて、評価するものであり、抵抗係数は、円柱に対する細長物体理論から求められる。鞭毛による推力と球に対するStokesの抵抗則で評価する細胞に作用する抗力を釣合わせて推進速度が求められる。このように抵抗力理論は、簡便ではあるが、あくまで近似解法であり、鞭毛の抵抗係数の選択にあいまいさが残り、定量的な結果には限界がある。一方、細菌も鞭毛を用いて運動するが、精子の鞭毛のように構造を持ち屈曲するものでなく、螺旋形の高分子重合体が菌体のべん毛モーターによって回転することによって、水中を運動する。そのため、細菌の場合を、べん毛と記すことによって区別することがある。Kellerら(5)は、細菌を、球形の菌体と1本の螺旋曲線のべん毛からなるとモデル化し、抵抗力理論を適用した。回転するべん毛によるトルクを打ち消すために、菌体が逆回転し、その結果、細菌は螺旋軌道を描きながら推進することを示した。このような鞭毛の研究に関する展望が、Lighthill(6)や、Brennenら(7)によって行われた。
微生物の運動を厳密に解析する努力が続けられ、Higdonは、鞭毛表面の境界条件を満足させるように、Stokesletとdipoletを鞭毛の中心に並べる細長物体理論を適用し、球形の細胞と屈曲する鞭毛からなる精子モデルおよび球形の菌体と螺旋型べん毛からなる細菌モデルに対する解析を行った(8)(9)。この細長物体理論では、正確解との誤差評価が可能である。また、Phan-Thienらは、境界要素法(BEM)を細菌の解析に適用した。BEMは球形以外の形状の菌体や複数のべん毛からなる細菌を対象にでき、境界要素を細かく取れば、正確な結果を得ることができる。
図1 精子の概略図
抵抗力理論の問題点として、抵抗係数の不正確さ並びに細胞の周りに発達する流れと鞭毛の相互作用が考慮されていないことが上げられる。Jonesら(11)は、細胞と鞭毛の相互作用を考慮できる抵抗力理論によって、クラミドモナスの運動を調べている。クラミドモナスは、数10μmのサイズのプランクトンであり、池などに生息し、生物対流を形成することでも知られている。図2に示すように、2本の鞭毛が細胞に着いており、平泳ぎのように運動する。この解析では、鞭毛が比較的短いことから、細胞の並進、回転運動によって形成される細胞周りのStokes流に対し鞭毛が相対運動するとして抵抗力の計算が行われている。国内では、小林ら(12)が鞭毛運動による推進機構に関する研究を進めている。谷ら(13)による鞭毛の屈曲運動の生物学的研究においては、鞭毛に作用する応力を抵抗力理論に基づいて評価している。一方、大腸菌、サルモネラ菌、ビブリオ菌のべん毛の回転数の測定が行われ、抵抗力理論と比較されている(14)(15)。後藤ら(16)も、単毛性細菌の運動に対しBEMを適用した解析を行っている。また、相沢(17)による細菌のべん毛モーターの研究に関する著書も面白い。水中を運動する微生物の抵抗力を利用するメカニズムや、その解析手法は、今後開発される水中を自走するマイクロマシンの技術に利用できるものと考えられる。
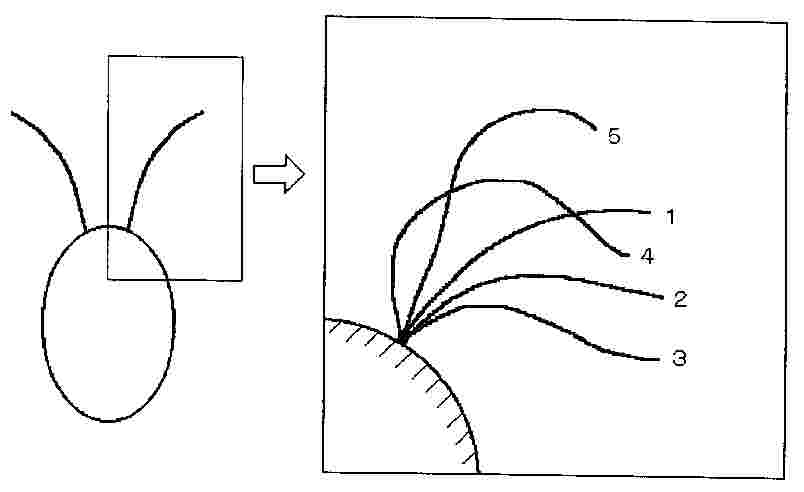
図2 クラミドモナスの動き
3.魚の推進機構(反動力推進)
レイノルズ数が高い遊泳速度の魚の推進機構は、Lighthill(18)の論文が基礎になっている。ほぼ同じ時期に、Wu(19)も、同様の研究を行った。いずれも、非粘性流体に対する細長物体理論あるいは薄翼理論を応用したものである。
水中を加速度運動する物体に作用する力には、物体の質量に仮想質量(付加質量)を加えて考えなければならない。これは、物体の運動に伴いその周りに流れが誘起されるために起こる。図3の魚の周りの流れの概略図に示すように、楕円柱をその長径に垂直方向に加速する場合、仮想質量は、長径を直径とする円柱の体積と等しい水の質量となる。魚が、体をくねらせて、反動力によって前向きの力を得るためには、表面の加速度の方向が後方成分を持つ必要がある。体全体に、くねらせる波動を後方に伝播させる場合を、ウナギ型推進と言う。また、体の後半3分の1程度を変動させる場合を、アジ型推進と呼ぶ。アジ型の魚の体は尾ひれの前で一旦くびれ、尾ひれでスパンが広くなる。尾ひれの効果は2次元の振動翼理論によって考慮されている。振動翼の周りに発達する循環は時間的に変化し、Kelvinの渦法則から空間全体の循環は保存されるので、図3に示すように、尾ひれの背後には、ひれの周りの循環の変化をキャンセルできるだけの渦が後縁から放出され、振動翼の場合のように、符号の異なる渦が次々と放出される。Chopraら(20)は、月形尾ひれの3次元形状を考慮した解析を行った。また、3次元振動板の解析が、Chengら(21)により、渦パネル法を用いて行われている。
最近の国内の研究について述べる。はじめに、魚の運動メカニズムなどの解説については、土屋(22)のものがある。最近、田中と永井(23)が水棲動物の遊泳の理論の解説と歴史および応用について詳しくまとめている。また、東(24)の「生物の動きの事典」には、水中や空中の生物の運動のメカニズムが網羅されており、水棲生物の運動や遊泳の理論が要領よくまとめられ、有用な式が集められている。中島ら(25)は、イルカ形水中推進機構について一連の解析を行っている。永井ら(26)は、振動翼の解析を行い、実験船を試作している。また、加藤ら(27)は、ブラックバスの泳ぎを観察し、胸ひれによる位置制御の解析を行い、魚型水中ロボットを開発している。
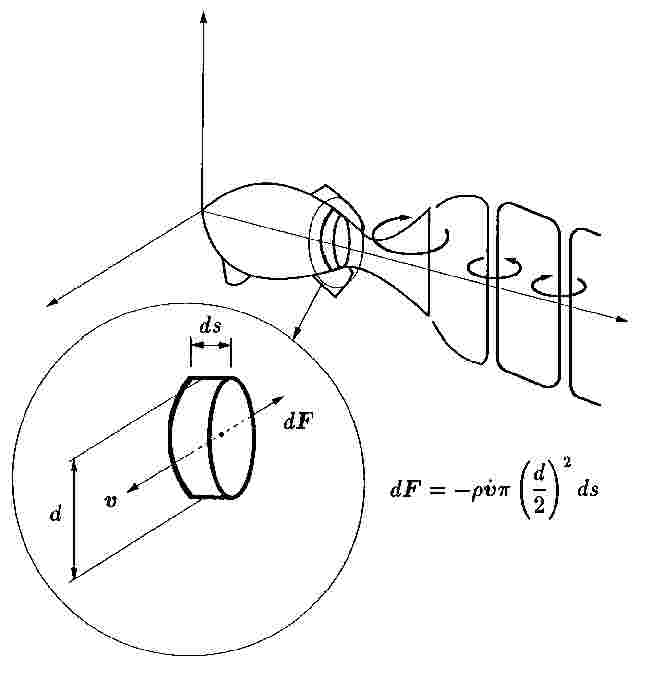
図3 魚の泳ぎと流れ
魚の周りの流れを把握するために、計算流体力学(CFD)による研究が行われている。蔦原ら(28)は、3次元離散渦法によって、Weis-Foghメカニズムの解析を行っている。蜂の空中静止飛行に関わるものであるが、尾ひれから放出される渦の数値解析にも、適用できるように思われる。秋本ら(29)は、差分法によって、屈曲変動する魚体の周りの流れ場の数値計算を行っている。Kimら(30)も、差分法により蠕動運動に誘起される流れ場の計算を行っている。また、劉ら(31)は、差分法による数値解析を行い、おたまじゃくしの泳ぎを調べている。差分法はNavier-Stokes方程式を直接数値計算するので、仮想質量の効果も含めて計算できる。しかし、計算結果を可視化する必要もあり、熟練を要するものである。最近、劉ら(32)は、差分法によって、昆虫の飛翔の3次元数値シミュレーションを行っている。
最近、水棲生物に関係するアクアバイオメカニズム研究会が発足しており、講演題目などは、ホーム・ページで参照できる。研究動向が、研究会の幹事である加藤(33)によってまとめられている。また、第10回バイオエンジニアリング講演会(34)のバイオミメティクスのオーガナイズド・セッションに水棲生物に関係する多数の発表があった。
4.バイオミメティクスと人工生命
以上に述べた魚やイルカの推進機構の解析は、水中推進機構を開発することを意図して行われている。これまでに、振動翼推進船(35)や小型自動機械魚(36)および胸ひれのある水中ロボット(33)が試作された。
一方、人工生命(ALife)の分野では、自律的に遊泳する魚のリアルなコンピュータ・シミュレーションが行われている(37)。図4に示すように、人工魚(Artificial Fish)の知覚学習に、研究の主眼が置かれている。人工魚の骨格は、支柱とスプリング・アクチュエータと質量を持つ交点からなり、交点に作用する内力および流体による外力を受ける運動系に対し、運動方程式を解くことで運動が決定される。コントローラーによりスプリング・アクチュエーターを加減することで体を変形させ、前進、方向転換を行う。また、眼(カメラ)と脳(コンピュータ)を有しており、画像処理を行い、強化学習によって効率的な泳ぎ方を修得する。他の魚と群れるとか、ある種の魚に恐怖感を抱くなどの習性に従って、自律的に遊泳するに至る。
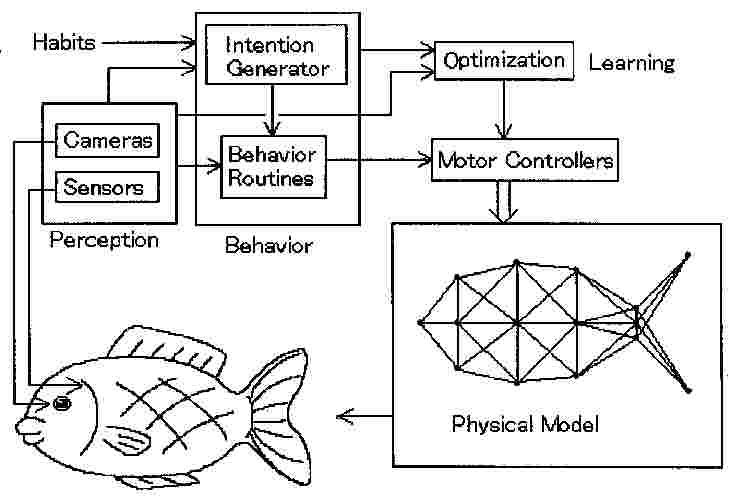
図4 人工魚(Artificial Fish)の知覚・学習および運動系の構成
Terzopoulosらの論文(37)では、流体力は表面速度と流速の相対速度に対する法線成分に比例すると仮定されている。魚の場合と同様、外力として、相対速度の時間微分に比例するとする反動力を仮定する方が現実的であろう。また、魚が、突然に方向転換する場合には、仮想質量による反動力と魚周りの循環および後流渦による揚力を合わせて考慮する必要があるので、大上ら(38)による加速する流れに対する特異点法を、魚のように変形する物体周りの流れに拡張する必要があろう。細菌は、べん毛を束ねて回転させることによって直線的に運動し、べん毛モーターを逆回転させることで束ねたべん毛を解き放ち方向転換を行う。このように直進と方向転換を繰り返しながら、細菌は誘引物質の回りに集まるという走化性を示す(17)。細菌は誘引物質の濃度変化を知覚でき、その濃度が上昇する限り、べん毛の回転を持続させ、濃度変化が低下する場合には、頻繁に方向転換することにより、目的地に接近できると考えられている。後藤ら(39)は、そのような簡単な行動規範に従って行動する細菌を模したマイクロ・ロボットの概念を提案し、そのロボットが目的地に接近できることをシミュレーションによって示した。
5.あとがき
これまでにも、流体力学の立場から水棲生物の遊泳に関心が持たれる時期が何回かあったが、最近、また、そのような研究が活発になってきたように思われる。一つは、大阪の海遊館を初めとする大型の水族館で、魚の遊泳をつぶさに観察できるようになったことがあり、水槽浄化装置が簡単に利用できるようになったためアーケードや割烹料理屋などでも水槽(いけす)で魚の泳ぎを見られる機会が増えてきたので、魚の運動に理解が深まり関心が持たれるためではないかと思っている。また、コンピュータと計算流体力学(CFD)の発達によって水棲生物の泳ぎと流れのシミュレーションが可能となったこともあろう。今後、多くの研究者がこの分野に参加されることを期待して、非学を省みず、水棲生物の研究についてまとめさせて頂いた次第である。
参考文献
1) Moffatt, K. : Nature, 394, p.728, (1998)
2) 神部 : ながれ, 17, p.383-385, (1998)
3) Lighthill, J : Mathematical Biofluiddynamics, SIAM (1975)
4) Gray, J & Hancock, GJ : J. Exp. Biol. 32, p.802-814, (1955)
5) Keller, JB & Rubinow, SI : Biophysical J. 16, p.151-170, (1976)
6) Lighthill, J : SIAM Review, 18, p.161-230 (1976)
7) Brennen, C & Winet H : Ann. Rev. Fluid Mech. 9, p.339-398 (1977)
8) Higdon, JJL : J. Fluid Mech. 94-4, p.685-711, (1979)
9) Higdon, JJL : J. Fluid Mech. 94-2, p.331-351, (1979)
10) Phan-Thien, N, Tran-Cong, T & Ramia, M : J. Fluid Mech.184, p.533-549, (1987)
11) Jones, MS, Le Baron, L & Pedley, TJ : J. Fluid Mech. 281, p.137-158, (1994)
12) 小林, 森川 : 機論(C編), 62-593, p.223-229 (1996)
13) Tani T, & Kaminuma, S : J. exp. Biology, 201, p.1493-1503, (1998)
14) Kudo, S, Magariyama, Y & Aizawa, S : Nature, 346-6285, p.677-680, (1990)
15) Magariyama, Y et al : Nature, 371-6500 p.752, (1994)
16) 後藤, 多賀, 高野 : 機論(B編), 63-605, p.188-193, (1997)
17) 相沢 : 原子が生命に転じるとき, カッパ・サイエンス, (1994)
18) Lighthill, J : J. Fluid Mech. 44-2, p.265-301, (1970)
19) Wu, TY : J. Fluid Mech. 46-2, p.337-355, (1971)
20) Chopra, MG & Kambe, T : J. Fluid Mech. 79-1, p.49-69, (1977)
21) Cheng, JY, Zhuang, LX & Tong, BG : J. Fluid Mech. 232, p.341-355, (1991)
22) 土屋 : 動物のメカニズム (加藤編),朝倉書店, p.32-44, (1980)
23) 田中, 永井 : 抵抗と推進の流体力学, シップ・アンド・オーシャン財団, (1996)
24) 東 : 生物の動きの事典, 朝倉書店, (1997)
25) 中島,小野 : 機論(B編), 62-600, p.3044-3051, (1996)
26) 永井ほか3名 : 機論(B編), 62-593, p.200-206, (1996)
27) 加藤, 稲葉 : 日本造船学会誌, 182, p.129-139, (1997)
28) 蔦原, 村上 : 日本航空宇宙学会, 45ー523, p.435-443, (1997)
29) Akimoto, H & Miyata, H : Proc. 5th Int. Symp. on CFD, Sendai, p.13-18, (1993)
30) Kim, MC, Mori, K, Doi, Y & Xu, Q : 日本造船学会論文集, 183, p.27-33, (1998)
31) Liu, H, Wassersug, RJ & Kawachi, K : J. Exp. Biology, 199, p.1245-1260, (1996)
32) Liu, H & Kawachi, K : J. Comput. Phys. 146, p.124-156, (1998)
33) 加藤 : 日本造船学会誌, 830, p.544-551, (1998)
34) 第10回バイオエンジニアリング講演会講演集, No.97-72, (1998)
35) 一色, 森川 : 日本造船学会誌, 642, p.2ー9, (1982)
36) Nagai, M, Maeda, G & Irabe, K : Advances in Bioengineering, ASME, p.56, (1982)
37) Terzopoulos, D, Rabie, T & Grzeszczuk, R : A-Life 5 Proceedings, Japan, p.313-320, (1996)
38) 大上, 赤松 : 機論(B編), 53-488, p.1224-1231, (1986)
39) 後藤, 伊藤, 高野 : 機論(C編), 64-624, p.3032-3038 (1998)
 著者プロフィール
著者プロフィール
高野泰斉(たかのやすなり), 1969年京都大学工学部機械工学科卒業, 1974年京都大学大学院博士課程修了, 同年より京都大学工学部助手, 1985年鳥取大学工学部助教授(共通講座), 1992年鳥取大学工学部機械工学科教授, 反応性気体力学の研究に従事,最近は,衝撃波治療,生物流体力学に関連する研究を行っている.
勤務先住所 〒680-0945 鳥取市湖山町南4-101